2. La tignoletta della vite
La tignoletta della Vite (Lobesia botrana Den. & Schiff.) o Verme dell’uva o Eudemide è un lepidottero (Ordine: Lepidoptera) appartenente alla famiglia dei tortricidi (Tortricidae) descritto per la prima volta in Austria da Denis e Schiffermüller nel 1776.
2.1. Distribuzione
È una specie tipicamente paleartica presente in tutto il Sud Europa, nel Nord Africa (Marocco, Algeria, Libia, Egitto) e dal Medio Oriente fino al Giappone.
In Europa si è diffusa alla fine del XIX secolo soprattutto nelle regioni viticole meridionali, poi si è espansa nell’Europa centrale dove nelle zone più calde convive con Eupoecilia ambiguella o Tignola della vite. Rispetto a quest’ultima però, L. botrana è distribuita in un’area più limitata.
2.2. Piante ospiti
La pianta ospite prediletta è la vite, ma è in realtà una specie molto polifaga, capace di vivere su molte piante tipiche dell’ambiente mediterraneo. Tra le più importanti si possono ricordare: Daphne gnidium (Thymelaeaceae) che è ritenuta essere l’ospite originario (Nuzzi e Triggiani, 1982), Arbutus tra cui il corbezzolo (Ericaceae), Mirtus, Ribes, Olea tra cui l’olivo, Ziziphus tra cui il giuggiolo (Rhamnaceae), Clematis, Corpus e Ligustrum, anche se non di tutte è conosciuto il ruolo che esercitano sul vigore, la fecondità, la velocità di sviluppo degli individui (Balachowsky 1966).

Figura 1: Lobesia botrana (Zangheri 1992);

Figura 2:
Larva ed adulto (Informatore agrario (20) 1999). Larve und Imago
2.3. Descrizione dell’insetto
L’adulto, di dimensione variabile tra 8 e 11 mm di apertura alare ed una lunghezza a riposo di 6-7 mm, presenta tutte le caratteristiche tipiche dei tortricidi. Le ali anteriori sono grigiastre variegate di giallo brunastro, con qualche lieve sfumatura azzurrognola negli esemplari più intensamente colorati. Tra le varie macchiette sulle ali, la più vistosa parte dal bordo costale e si restringe verso quello anale.
Le tibie sono chiare e munite di spine apicali.
Gli adulti sono facilmente riconoscibili a vista e la determinazione non costringe ad analisi dei genitali.
Il dimorfismo sessuale non è marcato: i maschi, come per buona parte dei lepidotteri, hanno l’addome più sottile e terminante a punta, mentre nelle femmine, al microscopio, l’addome si presenta con una cavità in corrispondenza dell’ovopositore.
Le uova, di forma lenticolari (07x0,6mm), vengono deposte in prevalenza sui bottoni fiorali o sugli acini, meno frequentemente su bratteole, pedicelli, rachidi. Esse hanno dapprima colore bianco giallastro, poi imbruniscono da una parte nello stadio di “testa nera” e infine imbruniscono completamente prima della schiusa.
Le larve, che sono alla nascita di 0.95-0.98 mm, raggiungono di solito i 9-10 mm di lunghezza, passano da un color biancastro con capo scuro del primo stadio, a nocciola scuro ocraceo fino ad un verde brunastro con capo leggermente più chiaro degli stadi più avanzati. Le pseudozampe addominali e anali sono munite di uncini rispettivamente in numero di 35 e 25. Il pettine anale può avere anche 8 denti. (Tremblay 1981).
Le crisalidi hanno colorazione verdastra prima, bruna dopo qualche giorno e dimensioni normalmente maggiori per le femmine (4-6 mm). Il cono anale ha una terminazione a forma di ventaglio (cremaster) che presenta 4 setole dorsali e 4 medio-dorsali che distingue L. botrana da E. ambiguella (S. Zangheri ed al. 1992).

Figura 3: Larve di seconda generazione; (Zangheri 1992). Larven der zweiten Generation;
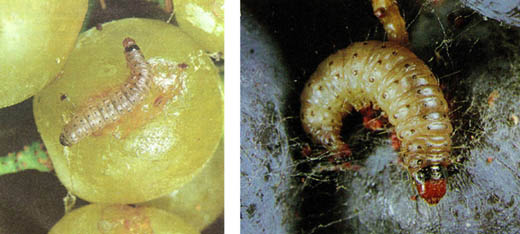
Figura 4: Larva di terza generazione (a destra, Ferrari 94, a sinistra Zangheri 1992). Larve der dritten Generation (rechts Ferrari 94, links Zangheri 1992)
2.4. Ciclo biologico della tignoletta della vite: Lobesia botrana.
Nei nostri ambienti la tignoletta si evolve con due-tre generazioni l’anno. Normalmente il picco di sfarfallamento della prima generazione si osserva nella prima decade di maggio, con massime presenze con il raggiungimento di 140 gradi-giorno (sommatoria dei valori di temperatura media giornaliere che dall'inizio dell'anno, superano il limite biologico di 10°C), anche se una parte degli sfarfallamenti è scalare e continua per tutto il mese.
I1 picco della seconda generazione si verifica di
regola a 7‑10 giorni dal principio delle catture che si hanno nella prima
decade di luglio, a volte qualche giorno dopo rispetto alla tignola.
Le femmine depongono in media 40-60 uova, disposte separatamente sugli acini, e le larve si schiudono dopo 4-7 gg. di sviluppo embrionale. Dai primi di agosto alla metà di settembre avviene il terzo volo dell’annata solitamente parziale. Le larve attaccano in Trentino i grappolini delle femminelle senza quindi comportare danni alla produzione.
Gli adulti hanno abitudini crepuscolari e notturne e sono particolarmente attivi con temperature superiori a 15°C. L’accoppiamento, la ovideposizione avvengono appunto nelle due ore successive al tramonto del sole, con temperature tra i 15° ed i 25°C e con una illuminazione compresa fra valori di 60-80 lux (Chaboussou, 1962). Entro alcuni giorni avvengono gli accoppiamenti e le femmine depongono mediamente una cinquantina di uova incollandole preferibilmente sulle brattee dei grappolini fiorali, meno frequentemente sul rachide e sui pedicelli. Lo sviluppo embrionale è possibile con temperature superiori a 9°C. Le larve nascono al raggiungimento di 75 gradi-giorno, normalmente dopo un periodo d'incubazione che dura 8-12 giorni. Le larve della prima generazione si possono osservare sui grappoli dalla seconda metà di maggio a tutto giugno (prefioritura, fioritura e allegagione) e danneggiano i bottoni fiorali e i fiori, per completare il loro sviluppo in piena fioritura o poco più tardi. Le larve riuniscono con fili di seta gruppi di fiori e raggiunta la maturità, si incrisalidano all’interno.

Figura 5: Crisalide svernante; Überwinternde Puppe
Quelle della seconda generazione si rinvengono da metà luglio fino alla fine di agosto, a cavallo cioè delle fasi fenologiche che vanno dalla chiusura grappolo all’invaiatura completata, entrando in corrispondenza del punto di contatto tra due acini, mentre quelle di un’eventuale ed incompleta terza generazione, dall'inizio di settembre sino ad ottobre, attaccano gli acini dei grappolini delle femminelle. In ogni caso raggiunta la maturità le larve escono e si impupano all’interno di ricoveri sericei biancastri più o meno lassi costruiti all’interno del grappolo.
Sono le larve di seconda generazione che, raggiunta la maturità, cercano un riparo sotto il ritidoma del ceppo, nelle screpolature dei pali tutori in legno o in altri ricoveri, dove incrisalidano all’interno di un bozzoletto di seta. La diapausa invernale interessa in pratica gia alcune larve mature nella seconde quindicina di agosto e tutte le larve sviluppate in settembre. Essa è stimolata, già allo stadio di uovo e di larva di prima età dalla riduzione del fotoperiodo (<15ore luce), mentre viene poco influenzata dalla temperatura. (Deseö 1988 & Roehrich 1969).

Figura 6: Uova di tignoletta stadio di testa nera (Informatore agrario (20) 1999). Eier des Bekreuzten Traubenwicklers Schwarzkopfstadium (links Informatore Agrario (20) 1999).
2.5. Influenza dei fattori biotici e abiotici
La dinamica di popolazione di questo temuto insetto è regolata dall’azione combinata di fattori biotici e abiotici. Difficile è distinguere e valutare il peso degli uni rispetto agli altri nelle differenti condizioni agro-ecologiche. Si può affermare tuttavia che i fattori biotici, rappresentati da predatori, parassitoidi e patogeni, giocano un ruolo significativo nel limitare la densità della popolazione della tignoletta e ridurre considerevolmente i danni da essa arrecati alla vite.
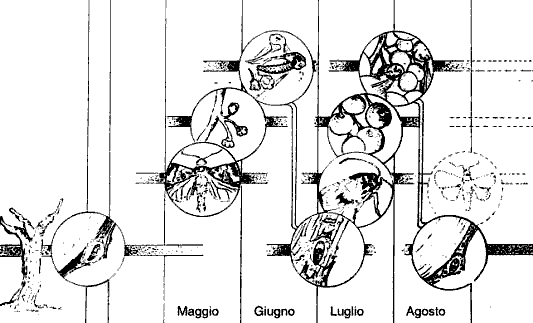
Figura 7: Ciclo biologico di Lobesia botrana (Baggiolini 1992). Lebenszyklus der Lobesia botrana (Baggiolini 1992)
2.5.1. Azione dei fattori abiotici
L’umidità relativa dell'aria esercita una forte influenza sull’attività degli adulti: alle temperature qui sopra indicate il volo è più intenso quando l'umidità relativa varia fra 40 e 70%. Questo caratterizza il differente comportamento di L. botrana e di E. ambiguella nei confronti dei fattori climatici. Nella Figura 8 si può notare come la parte tratteggiata corrisponda alla zona di attività per la prima e di inattività per la seconda (da Stellwaag, 1940).
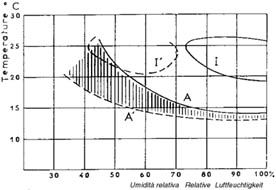
Figura 8: Ecoclimatogramma delle
esigenze ecologiche degli adulti di L.
botrana e E. ambiguella;
(Balachowsky, 1966). Ökoclimatogramm der
ökologischen Bedürfnisse der L. botrana
und E. ambiguella (Balachowsky, 1966).
A: limite della zona di
attività degli adulti di E. ambiguella;
Grenze der Aktivitätszone der adulten E. ambiguella;
A': limite della zona di
attività degli adulti di L. botrana;
Grenze der Aktivitätszone der adulten L. botrana;
1: zona di attività e
massima ovideposizione di E. ambiguella;
Aktivitätszone und maximale Eiablage der E. ambiguella;
1': zona di attività e
massima ovideposizione di L. botrana;
Aktivitätszone und maximale Eiablage der L. botrana;
La longevità delle femmine è strettamente correlata
con le condizioni ambientali, tuttavia la maggior parte delle uova sono deposte
nella prima settimana.
Studi sull’influenza della temperatura e
dell’umidità relativa dell'aria sullo sviluppo embrionale dell'uovo hanno
rilevato che la soglia termica, per lo sviluppo embrionale si colloca a circa 9°C, ma la schiusura esige una temperatura
leggermente superiore.
La durata di incubazione di uova poste in
termostato con umidità relativa costante (70%) è riportata nella Tabella 1.
|
°C |
Giorni/ Tag. |
|
34.5 |
4 |
|
29-33 |
3½ |
|
26 |
4½ |
|
24 |
4½ |
|
22 |
5½ |
|
17 |
10 |
|
20 |
7 |
|
15 |
14½ |
|
10.7 |
39 |
|
10.9 |
40 |
Tabella 1: Durata di incubazione
delle uova con umidità relativa costante. Inkubationsdauer
der Eier mit konstanter relativer Luftfeuchtigkeit (Götz 1940)
L’azione combinata di questi fattori porta a
differenti valori di schiusura delle uova in termini percentuali. Mentre a
temperature medie il tasso di mortalità è basso anche entro ampie oscillazioni
di umidità relativa, al di sopra di 22°C e al di sotto di 18°C la mortalità è
elevata con umidità relativa ridotta. (Götz, 1940).
La specie sembra inoltre ostacolata dal susseguirsi
di piogge che possono portare, come per tortricidi di altri fruttiferi, ad
annegamento della larva nel frutto.
2.5.2 Azione dei fattori
biotici
I fattori biotici sono stati oggetto di numerosi
studi tendenti a valutare la loro reale azione di contenimento o la possibilità
di una loro utilizzazione nella pratica.
Fra essi sono principalmente da considerare i
microrganismi patogeni, i predatori e i parassiti.
II ruolo dei predatori generici sembra importante,
anche se difficile da valutare. Si tratta, nella maggior parte dei casi, di
parassiti primari endofagi molto polifagi.
Lo stadio vitale maggiormente interessato dalla
parassitizzazione è quello di crisalide svernante e può raggiungere il 90% in
vigneti sufficientemente equilibrati, dove l’uso di fitofarmaci è oculato.
Un alto tasso di parassitizzazione invernale può
significare un ridotto attacco di prima generazione. Catoni infatti, suggeriva
di catturare con “fasce trappola” le crisalidi svernanti per liberare i
parassiti da esse sortiti.
È interessante notare, pur nella diversità della
ricerca e dei suoi obiettivi, i cambiamenti sopravvenuti nella composizione
faunistica ed il ruolo che le specie parassite esercitano nella limitazione
delle popolazioni di L. botrana. Le
specie sfarfallate da crisalidi svernanti di L. botrana ed E. ambiguella
in Trentino nel 1910, rilevate da Catoni, sono elencati nella Tabella 2.
È da notare la presenza predominante di Hymenoptera Ichneumonidae rispetto a Hymonoptera Chalcidoidea; attualmente
la situazione si è capovolta.
Le larve sono predate da Aracnidi, Dermatteri,
Neurotteri Crisopidi, Ditteri Sirfidi ed Imenotteri. Degni di nota, ma non
tutti presenti con popolazioni importanti in Trentino, sono la Dibrachys affinis, (Imenottero
Calcidoideo), che attacca anche le crisalidi, Eupelmus spp (Imenottero Calcidoideo).
Larve e crisalidi possono ancora essere attaccate
da funghi entomopatogeni quali Vertidihium
spp. e Poediomyces spp., nonché
da virus della poliedrosi citoplasmatica.
Tra i nemici, naturali il ruolo più importante è
sostenuto dai parassitoidi, dei quali sono state identificate più di 30 specie,
attive soprattutto su larve e crisalidi della terza generazione.
|
Famiglia/Familien |
Specie/Art |
|
ICHNEUMONIDAE |
Pimpla alternans |
|
Pimpla stringipleuris |
|
|
Pimpla detrita |
|
|
Angitia tenuipes |
|
|
|
Microcryptus nigrotincus |
|
|
Gambrus inferus |
|
|
Herpestomus furunculus |
|
|
Omorgus difformis |
|
|
Habrocytus puntiger |
|
|
Cinxaelotus eytrhrogaster |
|
|
Hemiteles areator |
|
CHALCIDOIDEA |
Eurytoma rosae |
|
|
Cricellis decipiens |
|
|
Habrocytus aculigona |
Tabella 2: Specie parassite di crisalidi svernanti di L. botrana ed E. ambiguella
in Trentino. (Catoni, 1910). Aus überwinternden
Puppen der L. botrana und der E. ambiguella
schlüpfende Parassitenarten im Trentino.
Tra i parassitoidi viene segnalata l’attività di
Imenotteri Calcidoidei (Eulofidi, Pteromalidi, Tricogrammatidi), Braconidi ed
Icneumonidi (Pimpla contemplator),
Ditteri Tanichidi (Phytornyptera
nigrina). Gli Icneumonidi (in particolare Dicaelotus inflerus, T. Campoplex
capitator e C. capita Aub.) sembrano avere un’importanza superiore
rispetto a quella degli altri parassitoidi. A fine inverno possono venire
raggiunte elevate percentuali di parassitizzazione ed alcuni di essi sono
ritenuti interessanti per i programmi di lotta biologica. (S. Zangheri 1992).

Tabella
3: Parassitoidi di
L. botrana rinvenuti nei vigneti
veneti. Parassiten der L.
botrana gefunden in den Weinbergen der Region Veneto. (Mescalchini e Della Monta 1998)
Dall’indagine sulla parassitizzazione della
tignoletta svolta dal laboratorio di difesa delle colture dell’Istituto Agrario
di San Michele all’Adige nel 1994 sono state rilevate le seguenti specie di
parassiti, espresse in percentuale rispetto al numero totale di crisalidi
raccolte.
|
Famiglia/Familien |
Specie/Art |
% |
|
ICHNEUMONIDAE |
Pimpla spuria |
6.34 |
|
|
Itoplectis alternans |
8.78 |
|
|
Dicelotus sp |
5.36 |
|
|
Triclistus sp |
4.39 |
|
|
Agrothereutes abbreviator |
0.49 |
|
|
Isadelphus sp |
0.49 |
|
CHALCIDOIDEA |
Dibrachys cavus |
2.44 |
|
|
Dibrachys affinis |
68.78 |
|
|
Eupelmus sp |
2.93 |
Tabella 4:
Percentuale di parassitizzazione delle crisalidi di Lobesia botrana. Prozentsatz an
Parassitierung der Puppen der L. botrana
L’azione letale dei parassitoidi si combina con
quella dei predatori e dei patogeni (funghi, protozoi e virus), in grado di
creare epidemie mortali sugli stadi giovanili.
Questo aspetto è evidente in modo particolare nei
vigneti non trattati con insetticidi di sintesi ma, pur giocando un ruolo
importante nel controllo biologico della tignoletta, non sono in grado di
limitare L. botrana in modo
significativo, senza che la loro preziosa attività venga salvaguardata mediante
l’adozione di razionali pratiche fitoiatriche.

Figura
9: Incidenza dei
diversi fattori di mortalità su larve e crisalidi di L. botrana in percentuali medie nei vigneti veneti. (Mescalchini e Della Monta 1998)
Einfluß der verschiedenen Mortalitätsfaktoren
auf Larven und Puppen von Lobesia
botrana in Weinbergen des Veneto (durchschnittliche Prozentwerte). (Mescalchini e Della Monta
1998)
4. La difesa
4.1. Metodi di lotta
La vite viene colpita da diverse malattie fungine
ma, tendenzialmente, da poche specie di insetti e acari. Nei nostri ambienti,
questi ultimi sono rappresentati principalmente da tignoletta (Lobesia botrana), cicalina della vite (Empoasca vitis) e ragno rosso (Panonychus ulmi) che, negli ultimi
anni, risulta solo raramente un problema perché ben contenuto dagli Acari
Fitoseidi. Le malattie crittogamiche invece, sono sempre le più pericolose ed
infatti peronospora, oidio e botrite si manifestano talora con un'aggressività
così elevata da ostacolare l'efficacia dei mezzi di contenimento adottati.
La lotta contro la tignoletta della vite può essere
effettuata con tecniche di lotta guidata, che si basano sul monitoraggio
eseguito con l’uso di trappole sessuali, oppure con tecniche di campionamento.
Il campionamento randomizzato su un centinaio di
grappoli deve essere eseguito nelle epoche che corrispondono alle due-tre
generazioni e deve essere ripetuto un paio di volte per determinare l’andamento
della generazione e il picco di massima presenza.
Le epoche corrispondono grossomodo alle fasi
fenologiche della fioritura per la prima generazione, al periodo che va dalla
mignolatura alla prechiusura grappolo per la seconda generazione e
dall’invaiatura in poi per la terza generazione, laddove questa ha luogo.
Nei confronti delle larve della prima generazione,
l’eventuale ricorso ad un intervento chimico può avvenire nella fase dei
bottoni fiorali separati con infestazioni medie su almeno il 35-40% dei grappoli per le varietà da vino con grappolo
spargolo o per varietà sensibili (Marzemino), e di 50% per quelle a grappolo
serrato (Chardonnay). Difficilmente comunque si raggiungono nelle nostre zone
attacchi così massicci per la prima generazione, che normalmente viene
monitorata quale parametro per la determinazione dell’epoca in cui avranno luogo le generazioni
carpofaghe.
Il volo della seconda generazione inizia
normalmente nella terza decade di giugno e si protrae fino alla seconda di
luglio.
Il volo si intensifica all’inizio di luglio (epoca
in cui si rinvengono le prime uova); i controlli per la determinazione del
momento del trattamento e dell’entità dell’attacco avvengono tramite l’utilizzo
di trappole sessuali e consistono nel monitorare le catture settimanali, onde
determinare il picco di massima presenza.
Questo si verifica normalmente con catture di 15-20
maschi per trappola per settimana, ma tale indicazione deve essere supportata
dal controllo dei grappoli, per rilevarne la percentuale con uova. Il
trattamento va effettuato con presenze di uova su almeno il 3-5% dei grappoli
per i vitigni pregiati e suscettibili agli attacchi di Botrytis cinerea (Pinot grigio, Pinot bianco, Pinot nero e
Chardonnay) e il 5-8% per gli altri.
Nei vigneti trentini è questa la generazione più
importante ai fini dei danni che possono essere provocati. La capsula
contenente il feromone ed eventualmente il fondo della tignoletta vanno
rinnovati circa 10-12 gg. prima del previsto volo della generazione successiva.

Figura 10:
Danno da tignoletta della prima generazione (Zangheri 1992). Schaden des Bekreuzten
Traubenwicklers der ersten Generation (Zangheri 1992)
Al superamento della soglia di intervento è
possibile eseguire dei trattamenti a base di insetticidi chimici o biologici.
Nel corso degli anni, l’evoluzione nella scelta dei
fitofarmaci ha risentito di molti studi che hanno permesso di valutare, oltre
all’efficacia di azione, la tossicità e gli effetti collaterali degli stessi.
I fosforganici con debole attività citotropica o a
lenta azione (fosalone, tebufenozide e formulazioni microcapsulate di esteri
fosforici) vanno applicati alla schiusa delle uova. I fosforganici
ovo-larvicidi e con attività citotropica (es. clorpyriphos-metil e quinalfos)
possono essere impiegati alla nascita delle larve o alla comparsa dei fori di
penetrazione.
Tra gli insetticidi biologici i più utilizzati sono
i preparati biologici a base di Bacillus
thuringiensis.
Bacillus
thuringiensis Berliner è un batterio gram-positivo, aerobio, sporigeno, flagellato,
reperibile nei terreni ed in diversi ambienti di tutto il pianeta, scoperto in
Giappone nei primi del ‘900. II primo formulato fu commercializzato in Francia
nel 1938, altri ne seguirono nell’immediato dopoguerra in diversi Paesi. La
sottospecie kurstaki, quella oggi
utilizzata, è stata scoperta nel 1962. Attualmente gli insetticidi a base di Bacillus thuringiensis rappresentano la
voce più importante del mercato dei mezzi di controllo biologico degli insetti,
minacciati in questo primato soltanto dal diffondersi delle piante
transgeniche, resistenti agli insetti, che peraltro esprimono nei propri
tessuti le tossine dello stesso microrganismo. (Caroli et Al. 1998)
La parte attiva è costituita dalla endospora o
corpo parasporale, una forma cristallina prodotta dal batterio, composta da
proteine di peso molecolare dai 27 ai 140 kDa altamente tossiche, delle quali
la più importante è la d-endotossina, che rappresenta il composto principale dell’insetticida.
Il corpo parasporale può avere origine da molti batteri e ha forma variabile in
base al tipo di battere che la produce. Le proteine del cristallo hanno una
dimensione che varia ed hanno il significato funzionale di protossine, dal
momento che è generalmente necessaria una loro parziale degradazione enzimatica
per liberare la frazione con attività insetticida.
A lungo si è creduto che lo spettro
di azione dei ceppi di Bacillus
thuringiensis fosse limitato alle specie dell’ordine dei Lepidotteri. A
partire dalla scoperta nel 1976 e nel 1982 di ceppi attivi, rispettivamente
contro Ditteri e Coleotteri, lo spettro di attività si è via via
allargato fino a comprendere attualmente anche Imenotteri, Rincoti e
Mallofagi,
Acari, nonché alcuni Nematelminti e Platelminti (Feitelson, 1993).
Nella continua ricerca di ceppi con nuove e
migliori caratteristiche insetticide e tecnologiche, i ricercatori di tutto il
mondo hanno collezionato, negli ultimi decenni, migliaia di isolati naturali
dai più svariati ambienti fra cui il terreno, polveri di cereali, insetti
infetti.
Il Bacillus
thuringiensis agisce quindi esclusivamente per ingestione esplicando la sua
azione tossica a livello intestinale. Applicazioni fogliari possono di
conseguenza raggiungere solo quei fitofagi con apparato boccale masticatore e
che si nutrono delle parti del vegetale esposto al trattamento (le d-endotossine
non hanno attività citotropica o sistemica).
Possono essere utilizzati sospensioni di Bacillus thuringiensis var. kurstaki (con l’eventuale aggiunta di
0,5 kg di zucchero per ettolitro d’acqua), da distribuirsi nell’imminenza della
schiusura delle uova, 9-10 giorni dopo l’impennata delle catture. Il risultato
si ottiene intervenendo entro due giorni dall’inizio della schiusura delle uova
e ripetendo il trattamento nel caso che la nascita delle larve si prolunghi
oltre sette giorni.

Figura
11: Danno da tignoletta con muffa grigia; Schaden des Bekreuzten Traubenwicklers mit Grauschimmel.
Nella prova eseguita nel 1988 dall’istituto Agrario
di San Michele all’Adige e dall’ESAT (Ente Servizio Assistenza Tecnica) su Cabernet e Teroldego, il trattamento è
stato effettuato immediatamente dopo il picco delle catture e rinnovato la
settimana successiva e si sono ottenuti riduzioni maggiori del 95%.
Nell'ambito della tignoletta della vite il
"sistema di lotta", negli anni, ha visto una fase di sostituzione dei
vecchi prodotti (metil parathion) con molecole dotate di una tossicità acuta ed
un impatto ambientale meno aggressivo (clorpirifos metil), fino a giungere a
molecole di origine naturale Bacillus
thuringiensis) e a metodi alternativi ai trattamenti veri
e propri: la confusione sessuale.

Figura
12: danno da
tignoletta di seconda e terza generazione

Figura 13:
Trappola sessuale. Pheromonfalle
5. Tecnica di lotta per
confusione sessuale.
L’evoluzione degli studi di base, parallelamente
all’esplorazione della chimica in settori nuovissimi (ormoni, metaboliti,
etc.), ha aperto la strada a quel gruppo di mezzi di protezione identificati
sotto il termine generale di mediatori chimici.
Ci è quindi stato possibile accedere al mondo degli
insetti, fatto di segnali chimici, onde comprendere come essi si riconoscono,
si riproducono e quali sono i rapporti che instaurano con le piante.
Pur dotati di un’ottima vista, gli insetti
utilizzano soprattutto l’odorato e il gusto per riconoscere il partner
sessuale, la preda o la pianta ospite. Vi sono pertanto sostanze chimiche,
presenti nell’ambiente, che, percepite dall’insetto, ne regolano il
comportamento, sia alimentare che riproduttivo.
Le speranze di un’inversione di tendenza dell’agricoltura,
nel senso ecologico, riposano anche sulla possibilità di utilizzare queste
molecole attive come nuove sostanze fitosanitarie, in grado di modificare
l’ambiente del fitofago, rendendolo ad esso inospitale. I progressi più
ragguardevoli sono stati finora ottenuti con l’individuazione, la sintesi e
l’utilizzazione di attrattivi sessuali o feromoni: composti della serie
alifatica con 10-18 atomi di carbonio (la maggior parte termina con un gruppo
funzionale ossigenato: acetato, alcool o aldeide, talvolta con uno o due legami
insaturi).
Le applicazioni di feromoni liberano sostanze che
non sono pericolose per gli insetti utili, ne per le piante, gli animali e
l’uomo, non sono tossiche, neppure per la stessa specie combattuta; la loro
efficacia non
si basa su proprietà venefiche, ma semplicemente sul disturbo della
comunicazione.
I composti rilasciati nell’aria in quantità minima,
ma enorme per l’insetto, si decompongono rapidamente in ossido di carbonio ed
acqua una volta raggiunto il terreno, non lasciando alcun residuo su nessuna
materia.
5.1.
Secrezione e percezione dei feromoni
Nelle femmine di Lepidotteri il feromone è prodotto
da strutture ghiandolari, che si aprono nella gronda dorsale che separa
l’ottavo e il nono segmento addominale.
L’emissione del feromone ha luogo nel corso della
serata (Mani et al., 1974) allorché
la femmina si mette in posizione di richiamo: innalza l’estremità dell’addome
scoprendo l’ovopositore, che inarca perpendicolarmente all’asse del corpo
(Fluri et al., 1974). La quantità
d’attrattivo contenuta nell’addome di una femmina vergine varia notevolmente in
funzione della specie. Secondo Charmillot (1982) una femmina di Cydia pomonella L. ne conterrebbe circa
10 mg., mentre una di Adoxophyes
reticulana F.v.R. ne disporrebbe di 250-400 mg.
Nei maschi gli organi chemiorecettori sono i
sensilli tricodeali localizzati sulle antenne. Lo stimolo è generato dal
complesso molecolare del feromone allorché raggiunge la parte sensibile, il
dendrite della cellula nervosa recettrice, passando attraverso i pori o le
fenditure della parete. Il sensillo contiene delle cellule recettrici
specifiche per ogni composto feromonale (Priesner, 1979). Il messaggio è
trasmesso sotto forma di impulso nervoso ad un centro motore e quindi al
cervello (Boeckh, 1976).
La distanza alla quale il maschio può percepire una
femmina è in funzione del tasso d’emissione di feromone, della diluizione
dell’attrattivo nell’aria e della soglia di percezione degli organi recettori
(Shorey, 1976). Il maschio che percepisce l’attrattivo intraprende un volo a
zig-zag per andare a posarsi a qualche centimetro dalla femmina.
A questo punto incomincia il corteggiamento che, se la femmina accetta, si
conclude con l’accoppiamento.
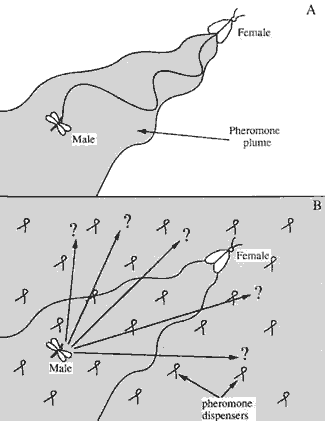
Figura 14:
A: Cono di feromone che guida il maschio alla femmina;
B:
Situazione resa indistinguibile dalla presenza di feromoni (Brunner J.F. 1991)
A: Pheromonkegel, welcher das Männchen zum
Weibchen lockt
B: Ununterscheidbare Situation aufgrund des
Vorhandensein der Pheromone
5.2. Storia dei feromoni
Risale agli anni ‘50-60
l’isolamento e la produzione del primo feromone attrattivo sessuale e fu il
bombicolo attivo sul Baco da seta Bombyx
mori. (Viggiani 1977)
Nel corso degli ultimi decenni i
progressi tecnologici effettuati dalla chimica analitica hanno
facilitato la messa a punto di
metodi che consentono l’isolamento,
l’identificazione e la sintesi di feromoni
d’insetti, aprendo così la via a nuove tecniche di difesa delle colture.
La prima esperienza di confusione
fu condotta da Burgess (1964) contro Limantria
dispar L. ed in seguito numerose furono le ricerche sull’applicabilità di
tale metodo contro una gran varietà di Lepidotteri dannosi alle colture
forestali, ai fruttiferi, alle colture erbacee ed alle derrate alimentari.
Nell’ultimo trentennio i feromoni sessuali di centinaia di insetti sono stati
identificati e sono state sintetizzate altrettante sostanze che, seppur
diverse, hanno la capacità di impedire ai maschi di trovare e fecondare le
femmine.
Nonostante ciò le sostanze oggi
sintetizzate non sono ancora state perfezionate e molte ricerche devono ancora
essere fatte perché i preparati che si utilizzano in confusione possiedano
tutte le caratteristiche di durata in conservazione, costanza di emissione e
stabilità agli agenti esterni necessarie per la riuscita della confusione.
In molti casi infatti, i feromoni
sono stati ottenuti con preparazioni incomplete, con miscele di prodotti chimici
attivi contro diverse specie, oppure con sostanze contenenti impurità, di cui non sempre si conosceva
l'effetto.
Sembra logico pensare che sostanze
non ancora perfezionate possano rilasciare un “buquet di odori”, che solo
marginalmente abbiano effetto di confusione e quindi necessitino di essere
applicate ad una concentrazione più alta, immettendo nell’ambiente anche
sostanze superflue.
Negli anni ‘70 Basf ha testato
ripetutamente il metodo della confusione sessuale, ma le prime esperienze
concrete in Italia furono fatte nel 1986 nella lotta alla Cydia molesta (C. Ioriatti, H. Arn, 1992).
Dalla fine degli anni ‘80 i
feromoni sono stati gradualmente introdotti nella pratica di campo in Trentino
come in altri distretti viticoli europei (Svizzera, Francia e Germania),
consolidando il metodo come una valida alternativa al sistema chimico.
La produzione delle miscele
feromonali segue in pratica tre vie possibili:
1)
la miscela ottimizzata: l'unione di varie sostanze che nel loro complesso
rappresentino l'approssimazione più vicina possibile al feromone emesso per la
femmina;
2)
un feromone incompleto: un componente del feromone ottenuto in modo più o meno
accurato che assomiglia quello della femmina;
3) una sostanza
analoga o in grado di mimetizzarsi, cioè la creazione di una sostanza che,
seppur diversa, è in grado alle condizioni di trattamento, di mimetizzarsi con
"la sostanza originale" e confondere i maschi;
Gli esperti non sono d'accordo su quale di queste
tre vie sia la migliore.
D'altra parte, risultati incoraggianti sono stati
ottenuti da regolari componenti del feromone con miscele. La controversia
continuerà probabilmente finché non si conosceranno perfettamente i meccanismi
dell'accoppiamento. (Henrich Arn,1992)
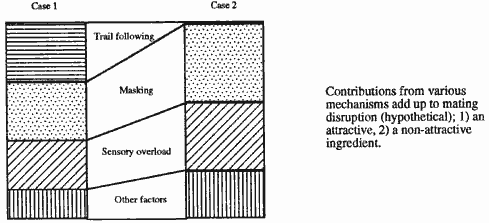
Figura 15:
Meccanismi di mascheramento del feromone femminile. (Harn 1992).
Überdeckungsmechanismen des weiblichen Pheromons. (Harn 1992)
Troppi sono i fattori che influenzano l’esito della
confusione per poter determinare gli elementi chiave che impediscono al maschio
di individuare le femmine.
Le ipotesi oggi maggiormente accreditate come
responsabili di successi ed insuccessi sono quattro:
1) l’inseguimento
dei maschi di una traccia falsa. Per concorrenza (trial following) fra varie
sorgenti di emissione (femmine e diffusori) (Roehrich e Carles, 1985) i maschi
sono attratti dalle fonti di odore artificiale che, competendo con quelle delle
femmine, fanno cambiare loro rotta troppo spesso.
2) l'odore delle
femmine viene mascherato dai feromoni. Il meccanismo di inibizione potrebbe
avvenire per mascheramento (mascking) del feromone naturale da parte di quello
sintetico (Neumann et al. 1986). I
maschi non possono più distinguere tra l'odore delle femmine e quello di fondo
creato dalle fonti artificiali.
3)
l’eccessivo carico sensoriale. Vi è una terza ipotesi che chiama in causa
l’eccessivo carico sensoriale (sensory overload). La stimolazione causata dai
feromoni sintetici provoca una riduzione della risposta agli stimoli sessuali a
livello del sistema nervoso periferico (adattamento) o di quello centrale
(abitudine, assuefazione) (Arn 1992).
Il contributo di ognuno di questi
meccanismi può dipendere dalla natura dell'ingrediente attivo, dalla sua
concentrazione e dalla sua distribuzione nello spazio e nel tempo.
L’inseguimento di una falsa
traccia, ad esempio, non può operare efficacemente con un feromone incompleto.
Esso dipende anche dagli andamenti dell’emissione dei dispenser che deve
mantenersi dentro un “range” ottimale. La capacità del feromone di mascherare
la posizione della femmina aumenta probabilmente con l’aumentare dell’intensità
di ogni fonte e con l'ammontare generale di sostanza per ettaro. L’eccessivo
carico sensoriale può essere efficace fuori della fase di risposta
dell’insetto, e perciò è l'unico meccanismo che può usufruire del vantaggio di
un sistema operante ventiquattro ore su ventiquattro.
La messa a punto di questo metodo
di lotta richiede l’adattamento della diffusione, che è regolata da fattori
chimici e fisici, alla biologia e all’etologia dell’insetto nelle condizioni
particolari della coltura da proteggere. Combinato ad altri mezzi di
previsione, aiuta a stabilire il momento ottimale per l’effettuazione di un
campionamento o di un trattamento, facilitando le decisioni dell’agricoltore.
5.3.
Formulazione dell’attrattivo - I
dispenser
La formulazione deve permettere di
liberare l’attrattivo in maniera lenta e graduale nel corso del tempo,
proteggendolo dalla degradazione e riducendone gli sprechi. Diverse
formulazioni a lenta liberazione (slow release) sono state messe a punto per
l’emissione di attrattivo sintetico.
I principali fattori che determinano l’emissione
dell’attrattivo sono:
1) la struttura del
supporto, la sua forma, le sue dimensioni, la concentrazione di prodotto, le
eventuali interazioni fisico-chimiche fra supporto e attrattivo, il peso
molecolare di questo, la lunghezza della catena, la presenza di doppi legami,
la presenza di gruppi funzionali, il numero e la posizione delle insaturazioni
ed infine l’isomeria (Bierl - Leonhardt, 1982; Daterman, 1982; Quisumbing e
Kydonieus, 1982);
2) fattori
ambientali come la temperatura il vento e in minor misura i raggi
ultravioletti, che possono favorire la degradazione chimica del prodotto e del
supporto, provocando così una perdita d’efficacia (Weatherston et al., 1985).
Agendo sull’insieme di questi fattori si tenta di
elaborare un dispositivo di diffusione efficace, capace di assicurare, almeno
durante le ore di volo, una concentrazione minima di attrattivo nell'aria, che
consenta la rottura delle comunicazioni fra i sessi.
Un importante passo avanti sulla determinazione
della quantità di feromone rilasciata nell’aria dai dispenser è stato lo
sviluppo di un mezzo di misurazione elettroantennografico.
Principali tipologie di
diffusori adottati nel corso degli anni
Microfibrille
e fibre capillari cave. Le microfibrille sono dei piccoli tubicini di qualche decina di mm di
diametro, generalmente di materiale plastico. Sono riempite di attrattivo e di
solvente e chiuse ad una delle due estremità. Le microfibrille, miscelate a
colla, sono distribuite nella coltura tramite una apposita apparecchiatura.
L’emissione dell’attrattivo avviene essenzialmente attraverso l’estremità
aperta, con una intensità assai rilevante in un primo tempo e che poi si riduce
e rimane in seguito ad un livello abbastanza costante. L’emissione è regolata
unicamente dal diametro dell’apertura, mentre la lunghezza della microfibrilla
determina la sua rimanenza (Weatherston et al., 1985).
Sono anche utilizzati dei tubi capillari di
plastica, chiusi alle due estremità. In questo caso l’attrattivo è contenuto
all’interno del tubo ed impregnato nelle pareti. In questi diffusori, che sono
distribuiti normalmente, l’emissione avviene attraverso le pareti.
Laminati plastici polistratificati. Si tratta di diffusori in
cui uno strato contenente il principio
attivo è situato fra altri due
strati con funzione di barriera, attraverso la quale avviene una emissione lenta. Sono
disponibili come nastri da applicare manualmente. Lo spessore e la natura degli
strati di plastica permette teoricamente di regolare la diffusione e la
persistenza (Quisumbing e Kydonieus 1982).
Micro e
macrocapsule. Una goccia di principio attivo è inglobata da un involucro polimerico.
Le capsule che si ottengono hanno un diametro variabile da qualche micron a
qualche millimetro, in funzione della tecnica di fabbricazione. Possono essere
preparate delle sospensioni in acqua e distribuite con un normale
polverizzatore. Le dimensioni delle capsule, lo spessore e la permeabilità
dell’involucro permettono teoricamente di regolare l’emissione.
Diffusori a
spaghetto.
Consistono in tubicini di materiale plastico, cavi internamente, con un’anima
in ferro e saldati alle due estremità. (Figura
16)
L’attrattivo miscelato al solvente viene fatto
scorrere all’interno di questi tubicini di alcune decine di metri di lunghezza,
fino a completa evaporazione del solvente e omogenea distribuzione del
principio attivo.
L’emissione avviene attraverso le pareti e le
estremità aperte. I tubicini sono tagliati alla lunghezza desiderata ed
applicati manualmente.

Figura 16: Dispenser Shin Etsu (Informatore Agrario (20) 1999).
Piastrine. Si tratta di erogatori
laminari di natura cellulosica (dimensioni 3x5 cm e 1,5 mm di spessore) impregnati con l’attrattivo, miscelato a
due stabilizzanti che fungono da protettori nei riguardi degli agenti ossidanti
chimici e fisici. Al fine di adeguare la velocità di diffusione alle esigenze
imposte dal complesso ambiente-fitofago, nelle diverse prove si sono variate le
quantità di attrattivo per unità di superficie, il numero di applicazioni per
stagione e di erogatori per ettaro, la quantità di attrattivo e di
stabilizzanti per erogatore ed alcune caratteristiche del supporto (Figura 17).
Ampolle. Da alcuni anni è stato
elaborato un sistema di erogazione, costituito da ampolle in plastica
contenenti la miscela di attrattivo e stabilizzanti. La natura e lo spessore
del contenitore, oltreché la quantità di prodotto contenuto in esso, sono i
fattori che intervengono nel caratterizzare la diffusione. (Figura 17)

Figura
17: Diffusori per la confusione
sessuale: a sinistra, diffusori Donegani; a destra Ampolle Basf. (Informatore
fitopatologico (4) 1992). Verteiler für die
Verwirrungsmethode: links Verteiler nach Donegani, rechts Ampullen von Basf. (Informatore
fitopatologico (4) 1992)
Esistono numerosi altri tipi di diffusori di
preparazione artigianale che presentano vantaggi e svantaggi diversi in
funzione del prodotto che si intende diffondere e della biologia del fitofago.
Si tratta in generale di impregnare o di includere il feromone nella massa di
un supporto, che può essere caucciù, plastica od altro materiale artificiale o
naturale. Largamente sperimentati sono
stati i tubi
in caucciù di
2-3 mm. di diametro esterno con la parete di 1 mm. di spessore
(Charmillot, 1980).
Ancora a livello sperimentale sono allo studio
formulati di feromoni in microcapsule, che possono essere impiegati come
normali fitofarmaci.
5.4. Fattori di
variabilità dell’emissione
La concentrazione dell’attrattivo sintetico
nell’atmosfera di una coltura dipende da numerosi fattori chimici, fisici e
meteorologici che ne influenzano l’emissione, la diffusione, la degradazione e
l’evaporazione. D’altro canto la rottura delle comunicazioni, presupposto per
la confusione, esige una concentrazione specifica minimale, che deve essere
assicurata durante tutto il periodo di attività sessuale dell’insetto.
La natura chimico-fisica del supporto di
diffusione, per la sua porosità, la sua permeabilità, le sue dimensioni, il suo
colore, la sua affinità con l’attrattivo, determina in gran parte la velocità di emissione del prodotto.
L’aumento del numero di atomi di carbonio della
sostanza da diffondere rallenta di molto la velocità di liberazione. Anche la
natura del gruppo funzionale riveste un ruolo importante; infatti per molecole
con la stessa lunghezza della catena, impregnate nel caucciù, gli alcoli
diffondono due o tre volte più rapidamente degli acetati (Butler e McDonough,
1981; Charmillot, 1980a; Bier Leonhardt 1982).
La velocità d’emissione dell’attrattivo aumenta
molto rapidamente con il crescere della
temperatura e della velocità del vento. L’influenza della luce sulla
degradazione dei prodotti è particolarmente temibile in presenza di doppi
legami.
In pieno campo il vento determina una rapida
liberazione dell’attrattivo mentre la massa vegetate ha effetto opposto e può
assorbire una parte d’attrattivo, che libererà in seguito (Wall et al. 1981).
Anche la forma e le dimensioni della parcella
intervengono nella determinazione della concentrazione di attrattivo
nell’atmosfera della coltura. Essa non è infatti omogenea, si accresce dal lato
sotto vento al lato opposto a mano a mano che l’aria, a contatto con i
diffusori, si carica d’attrattivo.
La concentrazione del prodotto varia anche in base
a fattori biologici, in particolare nelle colture ad alto fusto in quanto i
movimenti convettivi dell’aria sono ostacolati e modificati dalla vegetazione.
Durata del periodo di volo
La prima condizione da esaudire per soddisfare le
esigenze della lotta per confusione consiste nell’assicurare un’emissione soddisfacente
d’attrattivo durante tutto il periodo di volo dell’insetto e in particolare nei
momenti di attività sessuale. Un’emissione inferiore rispetto alla durata del
volo determina inevitabilmente degli insuccessi, mentre un prolungamento della
diffusione oltre il periodo di volo causa uno spreco di prodotto generalmente
assai costoso.
Le variazioni climatiche annuali modificano l’epoca
e la durata del volo, è quindi indispensabile un certo margine di sicurezza
nell’adattamento del diffusore alla biologia dell’insetto. È comunque
impensabile che un solo tipo di diffusore possa rispondere alle esigenze della
tecnica della confusione per qualsiasi fitofago, qualunque sia il suo
attrattivo e il suo ciclo biologico.
Ritmo di attività sessuale
Nel corso della giornata l’attività sessuale è
generalmente limitata, poiché nelle specie che rivestono per noi maggiore
interesse essa si svolge al crepuscolo, durante la notte o all’alba, vale a
dire in un momento in cui le basse temperature, limitano fortemente l’emissione
dell’attrattivo. Ne deriva che le alte temperature che si verificano
generalmente durante il giorno, causano uno spreco rilevante di prodotto, che
d’altra parte è difficilmente contenibile, se non con dispositivi tecnologici
complicati ed a costi proibitivi.
Mobilità del fitofago
La tecnica della confusione non esercita alcuna
protezione sulla vegetazione, ma mira a sopprimere gli accoppiamenti. Di
conseguenza deve essere applicata su tutta la zona dove si accoppiano e volano
gli adulti suscettibili di ovideporre nella coltura che si intende proteggere.
L’applicazione di questa tecnica sarà semplificata nel caso si debba lottare
contro insetti che migrano a breve distanza, mentre sarà inapplicabile per
quelli che migrano a lunga distanza. (la tignoletta compie modeste migrazioni e
variabili di 20-50m)
Densità della popolazione
Numerose prove di laboratorio e di pieno campo
hanno dimostrato che, per una determinata emissione d’attrattivo, l’efficacia
della confusione diminuisce all’aumentare della densità della popolazione. È
probabile che, in condizioni di elevata densità, i fitofagi si possono
ritrovare senza utilizzare la comunicazione feromonale, ma solo attraverso
movimenti casuali (Roehrich et al. 1979). In presenza di una elevata densità di
popolazione, l’efficacia può essere ancora mantenuta, ma richiede un aumento
molto rilevante della dose di prodotto (Hagstrum e Davis 1982. Charmillot et al., 1985). Sembra quindi vantaggioso introdurre la lotta per confusione in
presenza di basse densità di popolazione, mirando ad una regolazione del
fitofago nel lungo periodo. Se la popolazione di partenza è troppo elevata, è
consigliabile ridurla preliminarmente per mezzo di un trattamento classico
(Charmillot 1986; Charmillot e Bloesch 1987).
Scelta del composto
feromonale
Si sa che la maggior parte dei lepidotteri produce
un “bouquet” feromonale con diversi composti. I composti principali servono
essenzialmente a guidare i maschi in volo fino alla sorgente, mentre i composti
minori intervengono nelle ultime fasi che conducono all’accoppiamento (Baker e
Cardè 1979).
5.5. Grapamone
Con il termine grapamone si indica (E,Z)7-9
dodecadienylacetato, estere dell’acido acetico con alcol a lunga catena a 12
atomi di carbonio, che presenta due doppi legami alternati a configurazione
opposta.

Chemische Formel des Grapamone: (E,Z) –7,9- dodecadienylacetate
(E7,Z9-12Ac)
Il grapamone è stato approfonditamente studiato in
questi ultimi anni da Henrich Arn del quale sono allegate le pagine web
riguardanti i feromoni della tignoletta, da lui e da altri presi in
considerazione.
5.5.1 Studio del comportamento degli adulti
in confusione
Alcuni ricercatori hanno adottato diverse tecniche
per studiare il comportamento di Lobesia
botrana in ambienti posti in confusione sessuale al fine di stabilire:
1) densità delle popolazioni presenti;
2) limiti della diffusione del feromone dagli
erogatori.
Come molte specie di lepidotteri tortricidi, Lobesia botrana non è uniformemente
distribuita nell’ambiente viticolo.
Ciò è sicuramente legato alla variazione del
microclima, anche in aree relativamente omogenee.
La densità di popolazione, inoltre, è frutto
dell’effetto dei vari fattori biotici ed abiotici.
I primi, rappresentati dagli antagonisti naturali o
da fattori interni alla specie, possono rappresentare un importante fattore
di contenimento, soprattutto
per la generazione destinata allo svernamento (Roat 1993).
Ma soprattutto alcuni fattori del clima
(temperatura e pioggia in particolare) possono avere profonde ricadute sulla
dinamica di popolazione.
Nel vigneto in confusione gli strumenti più
frequentemente utilizzati per controllo delle popolazioni sono i seguenti:
1) controllo delle
catture nelle trappole a feromone. Se tale controllo rappresenta un utile
strumento per il monitoraggio delle popolazioni in vigneti non sottoposti a
confusione sessuale, nei vigneti in confusione la cattura di adulti nelle
trappole può essere un indice di incompleta efficacia del metodo;
2) esposizione e
ricattura di femmine vergini. Questa tecnica, largamente impiegata nelle prove
sperimentali; consiste nel disseminare in varie parti del vigneto femmine
vergini di Lobesia botrana “al
guinzaglio”, cioè con un filo incollato al protorace. La raccolta e la dissezione
dell’addome alla ricerca degli spermatofori ci potrà dire se la femmina è stata
o no fecondata. Oltre alla difficoltà di generalizzare questo metodo nel
controllo della qualità di diffusione, va anche precisato che la femmina,
confinata in piccoli spazi e con possibilità ridotta di movimento, rappresenta
in modo incompleto la “fitnes” della femmina libera nell’ambiente;
3) trappole
sovraccaricate. L’alimento è costituito da 77% di vino, 12% di succo di mela,
10 % di zucchero e 1% di acido acetico;
4) trappole alimentari per catturare le femmine
e verificarne la verginità
5) controlli
visuali sui danni. Il controllo dei nidi attivi prodotti dalle larve di prima
generazione sui grappoli fiorali è certamente il più semplice e sicuro:
semplice in quanto può essere svolto dal viticoltore con un modesto impegno
temporale; sicuro perché, come risulta dalle numerose esperienza condotte nelle
nostre ed in altre zone viticole europee, il livello di infestazione osservato
nei grappoli fiorali è predittivo di quale potrà essere il danno provocato
dalle larve di seconda generazione. Le numerose esperienze pratiche concordano
nel ritenere la soglia di infestazione del
5-8% sui grappoli fiorali come limite di tolleranza ammessa. Superata
tale soglia, la confusione va aiutata con un trattamento insetticida sulle
larve della seconda generazione.
5.5.2. Concentrazione di feromone nell’aria
e corrispondenti di L. botrana.
Malgrado la grande varietà di materiali usati nelle
prove eseguite dal Laboratorio di Difesa dell’Istituto Agrario di San Michele
all’Adige nel 1989, per la costruzione dei diffusori e dei diversi additivi
impiegati, qualche anno fa è stata stimata una quantità di feromone (grapamone)
minima, necessaria al mantenimento della confusione in condizioni di bassa
densità di popolazione 34,7 mg/ha/ora.
Nel grafico è rappresentata la diffusione oraria
dell’attrattivo e la diffusione dell’erogatore nelle prove eseguite a San
Michele all’Adige nel 1988 con diffusori Shin Etsu. La ditta dichiarava un
contenuto di attrattivo di 75 mg, meno della metà rispetto ai 172 mg degli
attuali dispenser. La quantità di sostanza attiva per ettaro invece era simile
a quella oggi consigliata poiché si è passati dai 1000 diffusori/ha di un tempo
ai 500 applicati nelle prove degli ultimi tre anni.
I metodi usati per stabilire il livello di
diffusione dei feromoni sono quelli per pesata e per estrazione della carica
residua nei diffusori attraverso la tecnica gascromatografica. Il metodo
consiste nella pesata di coppie di diffusori esposti all’inizio della stagione
e prelevati ad intervalli regolari di tempo.
La quantità di prodotto erogato viene in tal modo
stimato per differenza di pesata. Questa tecnica non è esente da difetti, primo
fra tutti la possibilità di non considerare alterazioni di tipo chimico della
molecola attiva.
Il metodo più sicuro rimane quello per
analisi chimica del componente attivo utilizzando la stessa procedura di
esposizione e di ritiro degli erogatori.

Figura 18:
Diffusione oraria dell’attrattivo
6. Le applicazioni
pratiche
6.1. Il caso della
Svizzera
In Svizzera la lotta per confusione è stata
omologata nel 1987 per combattere E.
ambiguella e da allora questa tecnica è stata impiegata ogni anno su
400-500 ha di viti nella Svizzera orientale, dove questa specie è la sola
presente. Nella maggior parte degli altri vigneti, dove c’è la presenza di L. botrana, si è fatto ricorso a
prodotti insetticidi fino al 1996, anno in cui è avvenuta l’omologazione dei
diffusori a base di feromoni attivi sulla tignoletta.
Nel 1996 i diffusori Rac 1+2 della Basf sono stati
applicati sia contro L. botrana che
contro Eupoecilia ambiguella su una
superficie di 463 ha (3,1 % della superficie viticola).
Per la prima generazione il tasso medio di attacco
sui 43.000 grappoli esaminati era dello 0,9%, ma localmente sui bordi,
raggiungeva anche il 9% di attacco.
In seconda generazione il tasso medio di attacco
sulle parcelle che avevano superato la soglia del 5-10 % ha permesso di
raggiungere un risultato finale soddisfacente ed una percentuale di danno
dell’1,4 %.
Nel 1997 la confusione sessuale contro le 2 tignole
della vite ha raggiunto un’espansione notevole di 1.200 ha (cioè l’8% della
superficie viticola svizzera).
Il campionamento fatto alla fine della prima
generazione metteva in evidenza un attacco medio molto basso, del 2,4 %; solo
in alcuni casi, cioè dove la tecnica veniva applicata per la prima volta o dove
la pressione del fitofago era elevata, la percentuale di attacco si elevava
oltre il 10 %.
Nella seconda generazione sono stati applicati
trattamenti localizzati solo sulle superfici che avevano evidenziato un attacco
compreso tra il 5 ed il 10 % (solo su 80 ettari, cioè sul 7% dei vigneti in
confusione) ed a fine stagione il tasso medio di attacco è stato dell’1,8 %.
L’emissione di attrattivo di L. botrana è avvenuto regolarmente ed a fine stagione le ampolle
contenevano ancora una quantità importante di feromone.
Al
contrario, per Eupoecilia
ambiguella l’emissione è
avvenuta rapidamente e in molti casi si è dovuto ricorrere ad un
apporto supplementare dei diffusori all’inizio del secondo volo.
Nel 1998 il ricorso alla tecnica di confusione ha
conosciuto una progressione spettacolare; è stato infatti adottato su 3.586 ha
(24% dei vigneti svizzeri). Nella maggior parte degli appezzamenti gli
erogatori Basf sono stati distribuiti con una densità di 500 unità per ha,
secondo quindi le indicazioni riportate; tuttavia in qualche vigneto in cui la
tecnica di confusione veniva applicata da più anni o dove le popolazioni erano
ridotte, la densità di diffusori è stata abbassata a 250-400 ampolle /ha.
I risultati ottenuti hanno confermato la validità
della tecnica, nonostante una pressione maggiore che negli anni precedenti e
soprattutto di Eupoecilia ambiguella.
In alcuni casi quindi si è dovuto fare ricorso ad
un trattamento in seconda generazione di quegli appezzamenti, in cui l’attacco
fiorale era superiore al 5-10%.
6.2. Il caso del Trentino
Nella Piana Rotaliana, dall’inizio degli anni ‘90,
si è attivato l’interesse ad applicare il metodo della confusione sessuale per
combattere la tignoletta. La vicinanza dei vigneti ai centri abitati, le
moderne tecniche di viticoltura, che richiedono una sempre maggiore e costante
presenza degli operatori in campagna, e la scelta di ottenere prodotti di
qualità, che si imponessero sui mercati più esigenti ha fatto sì che l’utilizzo
del metodo si espandesse anno dopo anno. Nel 1991 la prima area di 13,7 ha del
vigneto Sottodossi di Mezzocorona avviò l’esperienza; nel 1999 la lotta per
confusione sessuale ha riguardato ben 1.692 ha di vigneti, sia nella zona
pianeggiante che di altre zone del Trentino.
In base ai rilevamenti effettuati dalla ditta Shin
Etsu sui risultati della confusione sessuale, in Trentino dal 1992 al 1994 si
sono riscontrati attacchi molto bassi di tignoletta inferiori al 4 %.
Per l’esperienza sono stati utilizzati dispenser
ISONET-L con all’interno feromone (E,Z)-7, 9- dodecadienylacetato in dosi di
172 mg /dispenser, distribuiti in numero di 500/ha.
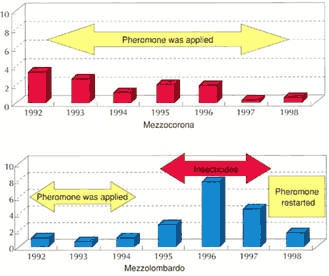
Figura 19: Andamento della prima generazione di tignoletta nell'ultimo sessennio
nella Piana Rotaliana. Entwicklung der ersten Generation des
Traubenwicklers während der letzten sechs Jahre im Piana Rotaliana.
Lungo i bordi della zona sottoposta al trattamento,
è stata fatta una barriera all’ingresso di femmine fecondate provenienti da
territori limitrofi, intensificando i diffusori.
II metodo si conferma molto efficace soprattutto
laddove siano rispettati due parametri fondamentali: vaste superfici coinvolte
e bassa densità della popolazione di tignoletta.
Dal 1995 l'applicazione è stata interrotta nella
zona di Mezzolombardo e mantenuta a Mezzocorona. Nel primo sito la popolazione
si è accresciuta molto dopo la sospensione del trattamento di confusione, come
indicato nella Figura
19.
Ulteriori esperienze sono in atto da quest’anno
sulla collina attorno a Trento, a Dro ed Arco in Valle del Sarca, ad Ala e
Rovereto in Vallagarina e a Telve in Valsugana.
7. Applicazione della
tecnica di confusione nell’azienda dell’Istituto
7.1. Piano sperimentale
La prova è stata condotta nell’azienda
dell’Istituto Agrario di San Michele all’Adige su una superficie di 23 ettari
di vigneto posto in collina ed allevato a pergola semplice trentina, con
diversi vitigni, come indicato nella Tabella 5.
Nella parte est i vigneti confinano con il bosco
ceduo, mentre sui restanti lati confinano con gli edifici e una strada
interpoderale che li separa dal frutteto.
7.2. Materiali e metodi.
Nella prova di confusione sessuale si sono
utilizzati diffusori composti da tubi in polietilene ad alta densità di colore
rosso (Figura 16) del diametro interno di 3 mm e quello esterno
di 5 mm impregnati con E7 Z9 –12ac,
(grapamone), il componente essenziale dell’attrattivo di Lobesia botrana.
Il diffusore utilizzato, prodotto dalla Shin Etsu,
è costituito da tubi capillari di plastica rinforzati da un filo di ferro e
riempiti con 172mg di feromone.
Al grapamone sono aggiunte due sostanze
stabilizzanti:
1,2 mg di BHT (2,6D terbutil-4methylfenolo) con
funzione antiossidante;
1,2 mg di sorbato (2,2 idroxy3
terbutil-5methyl-5clorobenzotriazolo) con funzione conservante.
Secondo quanto riportato in etichetta, i dispenser
sono stati applicati in numero di 500/ha (ogni spaghetto copre una superficie
di circa 20 mq.) attorcigliandoli una sola volta in modo lasco sui fili di
ferro o direttamente sui tralci ad un’altezza di 1,5 - 2 m.
La Shin Etsu dichiara per i diffusori una durata
tra i 150 e 200 giorni, secondo le condizioni di temperatura e vento presenti.
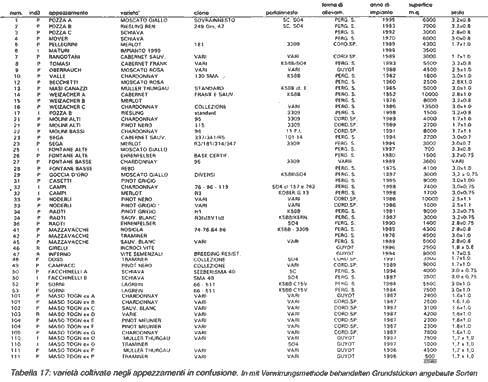
Tabella 5: varietà coltivate negli
appezzamenti in confusione
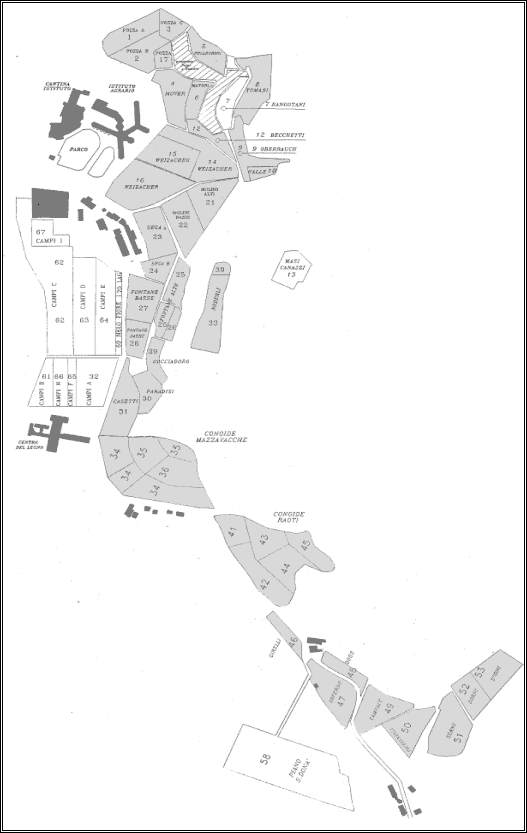
Figura 20:
Cartina dei vigneti trattati a confusione sessuale; Karte
der mit Verwirrungsmethode behandelten Grundstücke.
L’applicazione è stata eseguita nel modo più
regolare possibile nei giorni 22-24 aprile immediatamente prima dell’inizio del
volo degli adulti previsto con il calcolo della sommatoria termica e
controllato con le trappole sessuali a feromoni.
Nella fascia perimetrale del vigneto in confusione
sessuale (circa 30m) sono stati applicati un numero doppio di erogatori, al
fine di realizzare una barriera di protezione dalle immigrazioni.
I diffusori sono stati sottoposti ad una serie di
controlli di qualità, per verificare il corretto rilascio del feromone
nell’ambiente vigneto.
7.2.1. Controlli in campo
Sono stati effettuati i seguenti controlli:
- sulle trappole a feromone: Per verificare
l’efficacia di questo metodo è stato eseguito il controllo accurato di tre
trappole messe all’interno della zona interessata dalla confusione. L’assenza o
la presenza di maschi nelle trappole rappresenta uno dei parametri fondamentali
da considerare. Se il metodo funziona non si dovrebbero infatti registrare
delle catture.
- visuali sui grappoli fiorali e sugli acini:
Sono stati eseguiti dei controlli visuali atti a stimare l’attacco e la
quantità di uova.
Come
suggerito da Charmillot della Stazione Federale di Ricerche Agronomiche di
Changins nella Svizzera francese, il primo controllo è stato effettuato sulla
prima generazione per quantificare l’intensità della seconda generazione. Alla
fine di giugno si è quindi proceduto al rilevamento dei nidi delle larve e sono
stati osservati 500 grappolini fiorali per ogni appezzamento (per un totale di
20.000 grappoli).
Il secondo controllo mirava all’individuazione
dell’ovodeposizione della prima generazione. Sono stati osservati in maniera
randomizzata 200 grappoli per ogni appezzamento per un totale di 8.000
grappoli.
Il terzo controllo infine rappresentava
un’osservazione conclusiva per la verifica reale dell’efficacia del metodo. I tre controlli sono stati eseguiti per
fascia, separando il bordo dal centro del vigneto sottoposto a confusione
sessuale.
7.2.2. Controlli in
laboratorio
Sono state esposte varie coppie di diffusori per
l’analisi della quantità di feromone diffusa durante la stagione. Ogni 15
giorni dall’inizio dell’esposizione è stata prelevata una coppia di diffusori
per l’analisi dei residui di grapamone così da seguire l’evoluzione della
dispersione del prodotto.
I diffusori periodicamente prelevati sono stati
avvolti con carta stagnola per ridurre la perdita di feromone durante l’estate
e sono stati messi in freezer a –20°C.
Per le analisi si è proceduto all’estrazione del
grapamone secondo il seguente metodo:
I dispenser sono stati tagliati in diversi pezzetti
e lasciati per 24 ore immersi in 50 ml di esano. Successivamente sono stati
messi in bagno ad ultrasuoni per 15 minuti circa e sono infine stati lavati con
altri 50 ml di esano.
Si è proceduto quindi ad analisi per determinare la
quantità di sostanza attiva rimasta all’interno dei diffusori. È stato
utilizzato lo spettrofotometro di massa, il cui principio fisico consiste nella
possibilità di separare ioni atomici o molecolari con una o più cariche
positive per mezzo di un campo elettronico e di un campo magnetico,
opportunamente accoppiati.
Nello spettrofotometro di massa il campione viene
introdotto allo stato gassoso in una camera sotto vuoto, detta camera di
ionizzazione, dove la molecola viene bombardata con un fascio di elettroni, la
cui energia provoca la ionizzazione della molecola, con formazione dello ione
molecolare, la rottura dei legami, con frammentazione dello ione molecolare, ed
infine la ionizzanione dei frammenti.
Gli ioni così creati sono accelerati all’interno di
un tubo diritto, mediante un campo elettrico costante; ioni di massa diversa
acquistano ovviamente velocità differenti e giungono in tempi diversi al
sistema di rilevazione posto alla fine del condotto.
Le applicazioni della spettrofotometria di massa all’analisi
qualitativa, forniscono due tipi di informazioni: il peso molecolare e la
formula bruta dei composti in esame, e la struttura molecolare.
Il rilevatore genera un grafico dei picchi di
ricezione degli ioni, che giungono alla fine del tubo ed in base al numero di
questi picchi è possibile risalire alla sostanza analizzata.
7.3.
Risultati e discussione
7.3.1.
Risultati dei controlli in campo
Il volo della prima generazione di tignoletta è
avvenuto all’inizio di maggio ed ha manifestato la massima intensità degli
sfarfallamenti alla fine della prima decade del mese.
Il controllo sulla prima generazione ha evidenziato
un attacco massimo del 9-13% (come rappresentato nella tabella sottostante)
rilevato solo sui vitigni più sensibili siti in appezzamenti limitrofi al
bosco, quindi più esposti alle migrazioni.
|
Fuori confusione Außerhalb der Verwirrungszone |
Bordo (0-10 metri) Rand (0-10 Meter) |
Zona adiacente il bordo Dem Rande naheliegende Zone |
Centro Zentrum |
|
34.7 % |
13.1% |
9.1% |
2.5% |
Tabella 6:
Percentuale di grappoli con nidi di tignoletta. Prozentuelle Verteilung der Trauben mit
Traubenwicklernestern.
Dal secondo controllo, effettuato nella seconda
metà di luglio, si sono riscontrate presenze medie di uova su meno del 2% dei
grappoli. In seguito, con il controllo sulle larve della seconda generazione,
il danno è risultato limitato allo 0,9%, come indicato nella tabella
sottostante.
|
Bordo (0-10 metri) Rand (0-10 Meter) |
Centro Zentrum |
Danno medio ponderato Mittlerer Schaden |
|
1,7 |
0,5 |
0,9 |
Tabella 7:
Controllo finale, percentuale di
grappoli con larve Abschlußkontrolle, Prozent
der von Larven befallenen Trauben
Le catture di maschi di L. botrana registrate nelle tre trappole localizzate nei vigneti
sono state praticamente nulle nella trappola situata nella zona centrale,
mentre qualche esemplare (probabilmente casualmente attratto) è stato catturato
nelle due trappole poste nei bordi.
La bassa percentuale di danno medio
ponderato riscontrato su vigneti da più di tre anni trattati con il metodo
della confusione sessuale ribadisce ancora una volta che un approccio
continuativo del metodo permette di contenere entro livelli bassissimi
l’attacco di L. botrana.
Da altre prove, eseguite in Trentino in questi
ultimi anni, è stato inoltre possibile verificare, che già al primo anno di
applicazione la confusione sessuale riesce, senza l’ausilio di trattamenti
insetticidi, a limitare significativamente l’attacco delle generazioni
carpofaghe.
Ancora ridotta e per lo più limitata alle zone
centrali della zona trattata o in vigneti con basso livello di popolazione è
invece la possibilità di ridurre gli erogatori sotto il numero di 300/350
erogatori/ha.
Questi risultati, frutto di ormai un decennio di
prove e sperimentazioni, concorrono a consolidare ulteriormente questo metodo
di lotta, che si conferma come un sicuro sistema per combattere la tignoletta
della vite.
7.3.2
Risultati dell’analisi spettrofotometrica
Dalle analisi allo spettrofotometro di massa, sono
risultati i seguenti dati:
|
Codice della prova |
Data prelevamento |
Risultato |
|
D13599 |
15/04/99 |
171,73 |
|
D13600 |
03/05/99 |
171,39 |
|
D13601 |
17/05/99 |
149,53 |
|
D13602 |
01/06/99 |
133,88 |
|
D13603 |
15/06/99 |
125,52 |
|
D13604 |
01/07/99 |
110,74 |
|
D13605 |
15/07/99 |
81,55 |
|
D11907 |
05/08/99 |
81,72 |
|
D11900 |
18/08/99 |
64,76 |
|
D11903 |
06/09/99 |
43,50 |
|
D13606 |
21/10/99 |
29,78 |
Tabella 8:
Risultati dell’analisi spettrofotometrica sui diffusori. Ergebnisse
der spektrophotometrischen Analyse der Verteiler.
I dati ottenuti sono molto interessanti in quanto
delineano in maniera abbastanza chiara l’andamento della diffusione del
grapamone, che è stata tutto sommato regolare e sufficiente per confondere la L. botrana durante tutte le fasi di
sviluppo dei grappoli.

Figura 21:
Andamento della diffusione del grapamone. Diffusionsverlauf des Grapamone
Alla fine del presente
lavoro sono stati allegati i risultati della calibrazione dello
spettrofotometro e la scansione del quinto campione analizzato.
Il diffusore, prelevato il
21 ottobre e sottoposto ad analisi, ha consentito di valutare una presenza di
residuo pari a circa 30 mg/diffusore di grapamone; tale quantità è
probabilmente troppo bassa per rappresentare una riserva utile di sostanza
attiva, che possa essere presa in considerazione nell’annata successiva.
La diffusione oraria di
sostanza attiva emessa durante i 6 mesi di esperienze (15 settembre – 21
ottobre) è stata in media di 31,2 mg/ora,
pari a 15,6 g /ha/ora.
Tale quantità è circa la
metà di quella stimata qualche anno fa dalle prove sui dispenser Basf allora in
uso. Tale differenza è probabilmente da imputarsi al miglioramento dei materiali
dei diffusori e dei coformulati aggiunti al grapamone per migliorarne la
dispersione e la stabilità nel tempo.
8. Conclusioni
Nel corso degli ultimi 10 anni sono state raccolte
numerose osservazioni in seguito all’applicazione del metodo della confusione
sessuale. Il mondo agricolo incomincia a prendere confidenza con questa tecnica
di lotta e si sta sempre più diffondendo l’iniziativa di incrementare le
superfici interessate, proprio perché i principali benefici vengono assaporati
dai viticoltori stessi.
Nonostante la realtà viticola trentina sia
caratterizzata da un’elevata polverizzazione aziendale, (aspetto che
risulterebbe un grave limite per l’applicazione del metodo della confusione, se
non subentrasse l’attività delle strutture cooperative che coordinano l’elevato
numero di viticoltori), la tecnica di confusione è ormai diventata un sistema
abbastanza sicuro, tanto che già al primo anno può rappresentare un valido
sistema di lotta alla tignoletta della vite.
Ci sono però alcune considerazioni da tener
presenti: applicando questo metodo di contenimento della tignoletta e riducendo
così l’uso degli insetticidi può accadere che nuove problematiche possano
affermarsi nelle aree interessate. È infatti successo ad esempio nel 1998 che
in alcune zone, situate in una fascia compresa fra i 350 e i 450 m sulla
collina di Faedo, sono stati evidenziati danni da Eulia (Argyrotaenia pulchellana) fino ad un massimo dell’8% in alcuni
vigneti.
Questo insetto è molto polifago e compie tre
generazioni all’anno ma solo in realtà molto localizzate, la seconda
generazione può provocare dei danni sulla vite. La larva si ciba a spese del
rachide (con erosioni diffuse e forte produzione di seta rispetto alla
tignoletta) e degli acini. Generalmente i trattamenti sulla seconda generazione
di tignoletta contengono anche questo fitofago.
Da quando si applica
la confusione (anno 1991), il 1998 è il primo anno di comparsa di questo
fitofago; d’altro canto però è stato
osservato che l’applicazione del metodo della confusione unitamente ad una
riduzione della quantità e del tipo di fungicidi impiegati (solo Ziram, Delan e
IBS), comporta un arricchimento dell’agro-ecosistema soprattutto a favore delle
specie utili.
Gli effetti positivi sull’agroecosistema, a seguito
della riduzione dell’uso di insetticidi, si potranno osservare nei prossimi
anni.
Serve quindi ancora un vasto lavoro di indagine a
tanti livelli, ma positivo è il crescente interesse da parte delle strutture
cooperative e degli operatori; tutto questo ci consentirà un ulteriore
approfondimento delle conoscenze.
La viticoltura moderna richiede, infatti, una
presenza sempre più costante di operatori chiamati ad una serie crescente di
operazioni manuali. Poter lavorare in un ecosistema più pulito è perciò, prima
di tutto, un grande privilegio per gli addetti del settore.
Indice delle figure
Ringraziamenti
Voglio ringraziare tutti coloro che mi hanno
aiutato nella realizzazione di questo lavoro.
In particolare ringrazio:
- Tutti i componenti del Gruppo di Difesa
delle Colture dell’Istituto Agrario di San Michele all’Adige.
- Luisa Mattedi per la gentilezza dimostrata
e per il prezioso aiuto offertomi durante questi sei mesi di studio-ricerca.
- Claudio Rizzi per l’aiuto alla preparazione
dei dispenser all’analisi.
- Mirta Buzzi per le analisi
spettrofotometriche.
- Gino Angeli per la disponibilità, i
consigli e l’aiuto fornitomi durante questi tre anni di studio.
- Tutti i miei compagni di corso, in
particolare Reinhard Kerschbaumer, Wolfgang Tratter, Hannes Rottensteiner, Erwin
Eccli e Sabino Russo.
Bibliografia
Libri
1) Arn H. (1992). Mating
Distruption in orchards and vineyards, In:
Working group use of pheromones and other semiochemicals in integrated
control.
2) Baggiolini M. (1996), Osservazioni e ricerche
sulle tignole dell’uva nei vigneti ticinesi (Clysia ambiguella e Lobesia
Botrana).
3)
Baker T.C. e Carde R.T. (1979), Analysis of pheromone mediated behaviors in
male Grapholita molesta the oriental fruit moth (Lepidoptera Trortricidae),
Environ. Entomol. 8: 956-968.
4)
Bierl e Leonhardt B.A. (1982), Release netes from formulationend qualità
control methods, In: Kyodenieus
A.F., Beroza M., Insect suppression with controlled release pheromone systems,
CRC Press, Boca Raton Florida 1: 245-258.
5) Balachowsky
A.S., (1966), Entomologie appliquèe à l’agiuculture, Ed. Masson et Cie, Paris,
Volume primo.
6)
Boeckh J. (1976), Aspects of nervous codlingsensory quality in the factory
pathway of insect In: Proc. XV
Intern. Congr. Ent.: 308-322.
7)
Brunner J.F. (1991), Mating distruption as a control for fruit pests In: New directions in tree fruit pest
management.
8) Butler L.I. e McDonough L.M. (1981), Insect
sex pheromones: evaporation rates of alcohols end acetates from natural rubber
septa, J. Chem. Ecol.
7: 627-633.
9) Catoni G., (1910), Contributo per un metodo
pratico di difesa contro le tignole dell’uva, Stab. Tipografico ditta C.
Cassone, Casale Monf.
10) Chaboussou T.,
Carles J.P. (1962), Observatios sur le piégeage sexuel des mâles d’Eudémis,
Revue Zooligie agricole appl.7-9: 81-97.
11)
Daterman G.E. (1982), Monitoryng insects with pheromones: trapping objectives
end bait formulations In: Kydonieus
A.F., Beroza M., Insect suppression with controlled release pheromone systems,
CRC Press, Boca Raton Florida 1: 195-212.
12) Flaim L. (1998), Valutazione della resistenza
agli esteri fosforici nei lepidotteri tortricidi: studi su Lobesia botrana Den.& Schiff, Tesi di laurea
13) Fluri P., et al. (1974), Untersuchung über das
Paarungsverhalten des Apfelwicklers (Laspeyresia
pomonella L.) und über den Einfluss von künstlichern Sexuallockstoff auf
die Kopulationshäufigkeit, Mit. Schweiz. Ent. Ges. 47: 253-259.
14) Forti D., Ioriarri C., Mescalchin E., Mattedi
L. (1988), Prove di efficacia di fenoxycarb, Bacillus thuringiensis teflubenzuron
impiegati nella difesa contro la Tignoletta della vite. In: Atti giornate fitopatologiche, vol.2.
15) Götz B. (1940),
Lockflüssìgkeiten zur Beobactung des Traubenwi-cklersmottenfluges, Wein u. Rebe
22: 15-21.
16) Mani et al., (1974), Tagesrhythmus des Falterfangs
und Beobactungen über die Flugaktivitìät beim Apfelwichler (Laspeyresia pomonella L.), Mitt.
Schweiz. Ent. Ges. 47: 39-48.
17)
Neumann et al. (1986), E. ambiguella pheromone: the first
mathing distruption product for commercial use in german vineyards, Brittish
Crop protection Conference Pest end Desease: 371-375.
18) Pollini (1998), Manuale di entomologia
applicata, Edagricole, Bologna: 603-608.
19)
Priesner E. (1979), Specificity studies on pheromone receptor of Noctuid and
Tortricid Leodoptera. In: Ritter F.J., Chimical Enology:
Odour conunications in animals. Elsevier, North-Olland: 57-71
18) Roat Cinzia. (1990), La Tignoletta della Vite:
Studi di dinamica di popolazione, importanza e diffusione dei suoi nemici
naturali, Tesi di laurea.
19)
Quisumbing A.R. Kydonieus A.F. (1982), Laminated
structure dispensers. In: Kydonieus
A.F. Beroza M., Insect suppression with
controlled release pheromone systems. CRC. Press, Boca Raton, Florida. 1: 213-235.
20) Roehrich R.
(1969), La diapause de l’Eudemis de la vigne L. botrana. (Lép. Tortricidae): Induction et èlimination, Zool. Ecol. anim 1 : 419-431.
21) Rochrich R.,
Carles J. P., Tresor C. et al. (199 ), Essais de Confusion sexuelle contre les
Tortricides de la grappe l’Eudemis L.
botrana Den e Schiff et la Cochylis
ambiguella., Tr. Ann. Zool. Ecol. anim 11(4):659-675.
22) Roehrich R
Carles J.P. (1985), Essai de confusion contre Lobesia botrana en 1983 CR de lutte integrée en vignoble, O.I.L.B.
Srop, Bernkastel
23)
Shorey H H. (1976), Animal communication by pheromone romones, Acad Press: 166
24)
Staedler E. (1976), Sensory aspects of insects plant interactions. In: Proc. XV Intern. Congr. Ent.:
228-248.
25) Tremblay E. (1982), Entomologia applicata
generalità e mezzi di controllo. (85 Vol.1).
26) Viggiani G. (1977), Lotta biologica ed
integrata, Liguori editore.
27)
Weatherston L. Miller D. Lavoie-Dornik. J. (1985), Commercial hollow-fiber
pheromone formulations: the degrading effect of sunlight on celcon fibers
causing increased release rates of the active ingredient, Chem. Ecol. 11: 1631-1644
28) Zangheri et al. (1992), Lepidotteri die
fruttiferi e della vite, Bayer, Milano :81-103
Riviste
1) Boller E., Zuber
M. (1996), Verwirrung des Eibindighen und Breutzen Traubenwicklers, Schweiz. Z.
Obst-Weinbau 132 (24): 652-653.
2) Caroli, Valentini, Pasqualini (1998), Bacillus thuringiensis spp. kurstaki e aizawai Informatore fitopatologico. (7/8)
3) Charmillot P.J.
e Bloesch B. (1987), La technique de confusion sexuelle: un moyen spécifique de
lutte contre le carpocapse C. pomonella,
L. Revue Suisse Vitic. Arboric. hortic. 19 (2) :129-138.
4) Charmillot P.J.,
Pasquier D., Scalco A. (1995a), Six ans de lutte per confusion contre les vers
de la grappe eudèmis et cochylis avec une densitè rèduite de diffuseurs. Revue
Suisse Vitic. Arboric. Hortic. 27 (1) : 7-12
5) Charmillot P.J.,
Pasquier D., Scalco A. (1995b), Lutte par confusion contre les vers de la
grappe eudèmis et cochylis à Perroy. Rèsultats de 1994, Revue Suisse Vitic.
Arboric. Hortic. 27 (6) : 339-345.
6) Charmillot P.J.,
Pasquier D., Scalco A. (1995c), Lutte par confusion contre les vers de la
grappe eudèmis et cochylis à Perroy et Allaman. Rèsultats de 1995, Revue Suisse
Vitic. Arboric. Hortic. 27 (6): 347-358.
7) Charmillot P.J.,
Pasquier D., Schmid A. (1997), Lutte par confusion contre les vers de la grappe
eudèmis et cochylis en Suisse. Rèsultats de 1995, Revue Suisse Vitic. Arboric.
Hortic. 29 (5): 291-299.
8) Charmillot P.J.,
Pasquier D., Schmid A. (1997), Lutte par confusion en 1997 contre les vers de
la grappe eudèmis et cochylis en Suisse, Revue Suisse Vitic. Arboric. Hortic. 30 (3) : 201-206.
9)
Charmillot P.J., Pasquier D., Rac. (1999), Lutte par
confusion sexuelle contre les vers de la grappe bilan de l’annèe (1998), Revue
suisse Vitic. Arboric. Hortic. 31 (1): 12-13.
10) Dalla Montà L.
e Marchesini E. (1992), Rapport du groppe “Lutte intègrèe en viticulture“
O.I.L.B, Rèunion de Conegliano.
11) Forti
D., Ioriatti C,. (1987), I mediatori chimici nella protezione delle colture agrarie,
Informatore fitopatologico (10): 31-36.
12)
Ioriatti C., Arn H. (1992), Use of pheromones and other semiochemicals in
integrate control.
13) Ioriatti C, Rizzi C. (1992), La tecnica della
confusione sessuale: quattro anni di esperienze nel controllo di carpocapsa e
Tortricidi ricamatori del melo in Trentino, Informatore fitopatologico (04):
49-56.
14) Karmin A. (1997), Pesticide Profiles.
15) Marchesini, Dalla Montà (1998), I nemici
naturali della tignoletta dell’uva nei vigneti del veneto, Informatore
fitopatologico (9)
16) Remund U.,
Boller E., Zuber M. (1996), Verwirrungstechnik des Eibindighen und Breutzen
Traubenwicklers in der Ostschweiz, Schweiz. Z. Obst-Weinbau 132 (8): 212-215.
17) Roat C., Forti D. (1994), Indagine sulla
parassitizzazione della tignoletta della vite in Trentino, Bollettino ISMA II, 1: 38-41
18)* Varner M., Mattedi L., et al. (1996),
Esperienza con il metodo della confusione sessuale in viticoltura, Terra
trentina. (3) 24-35
19)* Varner M., Mattedi L. (1996), 5 anni di lotta alla
tignoletta dell’uva con il metodo della confusione sessuale a Mezzocorona,
Terra trentina (2) 38-46.